Характерной чертой водной среды является ее подвижность, особенно в проточных, быстро текущих ручьях и реках. В морях и океанах наблюдаются приливы и отливы, мощные течения, штормы. В озерах вода перемещается под действием температуры и ветра.

Температурный режим. Отличается в воде, во-первых, меньшим притоком тепла, во-вторых, большей стабильностью, чем на суше. Часть тепловой энергии, поступающей на поверхность воды, отражается, часть расходуется на испарение. Изменение температуры в текущих водах следует за ее изменениями в окружающем воздухе, отличаясь меньшей амплитудой.
В целом же следует отметить, что амплитуда годовых колебании температуры в верхних слоях океана не более 10-15 °С в континентальных водах 30—35°С.
Глубокие слои воды отличаются постоянством температуры. В экваториальных водах среднегодовая температура поверхностных слоев составляет 26—27°С, в полярных — около 0°С и ниже. Исключением являются термальные источники, где температура поверхностного слоя достигает 85—93°С.
В воде как среде жизни, с одной стороны, существует довольно значительное разнообразие температурных условий, а с другой — термодинамические особенности водной среды, такие, как высокая удельная теплоемкость, большая теплопроводность и расширение при замерзании (при этом лед образуется лишь сверху, а основная же толща воды не промерзает), создают благоприятные условия для живых организмов.
Так, для зимовки многолетних гидрофитов в реках и озерах большое значение имеет вертикальное распределение температур подо льдом. Наиболее плотная и наименее холодная вода с температурой 4°С располагается в придонном слое, куда опускаются зимующие почки (турионы) роголистника, пузырчатки, водокраса и др., а также целые облиственные растения, такие, как ряска, элодея.
Утвердилось мнение, что погружение связано с накоплением крахмала и утяжелением растений. К весне крахмал превращается в растворимые сахара и жиры, что делает почки легче и обеспечивает возможность их всплытия.
Организмы в водоемах умеренных широт хорошо приспособлены к сезонным вертикальным перемещениям слоев воды, к весенней и осенней гомотермии, к летней и зимней стагнации. Поскольку температурный режим водоемов характеризуется большой стабильностью, среди гидробионтов в большей мере, чем среди организмов суши, распространена стенотермность.
Эвритермные виды встречаются главным образом в мелких континентальных водоемах и на литорали морей высоких и умеренных широт, где значительны суточные и сезонные колебания.
Плотность воды. Вода отличается от воздуха большей плотностью. В этом отношении она в 800 раз превосходит воздушную среду. Плотность дистиллированной воды при температуре 4 °С равна 1 г/см3. Плотность же природных вод, содержащих растворенные соли, может быть больше: до 1,35 г/см3. В среднем в водной толще на каждые 10м глубины давление возрастает на 1 атмосферу. Высокая плотность воды отражается на строении тела гидрофитов. Так, если у наземных растений хорошо развиты механические ткани, обеспечивающие прочность стволов и стеблей, расположение механических и проводящих тканей по периферии стебля создает конструкцию «трубы», хорошо противостоящую изломам и изгибам, то у гидрофитов механические ткани сильно редуцированы, так как растения поддерживаются самой водой. Механические элементы и проводящие пучки довольно часто сосредоточены в центре стебля или листового черешка, что придает способность изгибаться при движениях воды.
Погруженные гидрофиты обладают хорошей плавучестью, создаваемой специальными приспособлениями (воздушные мешки, вздутия). Так, листья лягушатника лежат на поверхности воды и под каждым листом имеют наполненный воздухом плавучий пузырь. Как крошечный спасательный жилет, пузырь позволяет листу плавать на поверхности воды. Воздушные камеры в стебле поддерживают растение в вертикальном положении и доставляют кислород корням.
Плавучесть также повышается с увеличением поверхности тела. Это хорошо видно у микроскопических планктонных водорослей. Различные выросты тела помогают им свободно «парить» в толще воды.
Организмы в водной среде распределены по всей ее толще. Например, в океанических впадинах животные обнаружены на глубинах свыше 10 000 м, переносят давление от нескольких до сотен атмосфер. Так, пресноводные обитатели (жуки-плавунцы, туфельки, сувойки и др.) в опытах выдерживают до 600 атмосфер. Голотурии рода Elpidia, черви Priapulus caudatus обитают от прибрежной зоны до ультраабиссали. Вместе с тем следует отметить, что многие обитатели морей и океанов относительно стенобатны и приурочены к определенным глубинам. Это относится в первую очередь к мелководным и глубоководным видам. Только на литорали обитают кольчатый червь пескожил Arenicola, моллюски — морские блюдечки (Patella). На больших глубинах при давлении не менее 400-500 атмосфер встречаются рыбы из группы удильщиков, головоногие моллюски, ракообразные, морские звезды, погонофоры и другие.
Плотность воды обеспечивает возможность животным организмам опираться на нее, что особенно важно для бесскелетных форм. Опорность среды служит условием парения в воде. Именно к этому образу жизни приспособлены многие гидробионты.
Световой режим. На водные организмы большое влияние оказывают световой режим и прозрачность воды. Интенсивность света в воде сильно ослаблена, так как часть падающей радиации отражается от поверхности воды, другая поглощается ее толщей. Ослабление света связано с прозрачностью воды. В океанах, например, с большой прозрачностью на глубину 140 м еще падает около 1 % радиации, а в небольших озерах с несколько замкнутой водой уже на глубину 2м — всего лишь десятые доли процента.
В связи с тем что лучи разных участков солнечного спектра неодинаково поглощаются водой, с глубиной изменяется и спектральный состав света, ослабляются красные лучи. Сине-зеленые лучи проникают на значительные глубины. Сгущающиеся с глубиной сумерки в океане имеют вначале зеленый, затем голубой, синий, сине-фиолетовый цвет, сменяясь в дальнейшем постоянным мраком. Соответственно сменяют друг друга с глубиной и живые организмы.
Так, растения, живущие на поверхности воды, не испытывают недостатка света, а погруженные и особенно глубоководные относят к «теневой флоре». Им приходится адаптироваться не только к недостатку света, но и к изменению его состава выработкой дополнительных пигментов. Это прослеживается на известной закономерности окраски у водорослей, обитающих на разных глубинах. В мелководных зонах, где растениям еще доступны красные лучи, которые в наибольшей степени поглощаются хлорофиллом, как правило, преобладают зеленые водоросли. В более глубоких зонах встречаются бурые водоросли, имеющие кроме хлорофилла бурые пигменты фикофеин, фукоксантин и др. Еще глубже обитают красные водоросли, содержащие пигмент фико-эритрин. Здесь четко прослеживается способность к улавливанию солнечных лучей с разной длиной волны. Данное явление получило название хроматической адаптации.
Глубоководные виды имеют ряд физических черт, свойственных теневым растениям. Среди них следует отметить низкую точку компенсации фотосинтеза (30-100 лк), «теневой характер» световой кривой фотосинтеза с низким плато насыщения, у водорослей, например, крупные размеры хроматофоров. Тогда как у поверхности и плавающих форм эти кривые более «светлого» типа.
Для использования слабого света в процессе фотосинтеза требуется увеличенная площадь ассимилирующих органов. Так, стрелолист (Sagittaria sagittifolia) формирует разные по форме листья при развитии на суше и в воде.
В наследственной программе закодирована возможность развития в том и другом направлении. «Пусковым механизмом» для развития «водных» форм листьев служит затенение, а не непосредственное действие воды.
Нередко листья водных растений, погруженные в воду, сильно рассечены на узкие нитевидные доли, как, например, у роголистника, урути, пузырчаток, или имеют тонкую просвечивающую пластинку — подводные листья кубышек, кувшинок, листья погруженных рдестов.
Данные черты характерны и для водорослей, таких, как нитчатые водоросли, рассеченные талломы харовых, тонкие прозрачные талломы многих глубоководных видов. Это дает возможность гидрофитам увеличить отношение площади тела к объему, а следовательно, развивать большую поверхность при сравнительно небольших затратах органической массы.
У частично погруженных в воду растений хорошо выражена гетерофилия, т. е. различие строения надводных и подводных листьев у одного и того же растения: Это хорошо просматривается у водного лютика разнолистного (рисунок 1) Надводные имеют черты, обычные для листьев надземных растений (дорзовентральное строение, хорошо развитые покровные ткани и устьичный аппарат), подводные — очень тонкие или рассеченные листовые пластинки. Гетерофилия отмечена также у кувшинок и кубышек, стрелолиста и других видов.
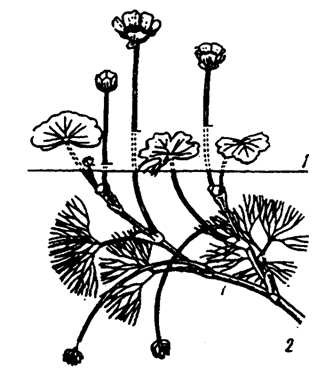
Рисунок 1 — Гетерофилия у водного лютика разнолистного Ranunculus diversifolius (из Т, Г. Горышиной, 1979) Листья: 1 — надводные; 2 — подводные
Показательным примером является поручейник (Simn latifolium), на стебле которого можно видеть несколько форм листьев, отражающих все переходы от типично наземных до типично водных.
Глубина водной среды оказывает влияние и на животных, их окраску, видовой состав и т. д. Например, в озерной экосистеме основная жизнь сосредоточена в слое воды, куда проникает количество света, достаточное для фотосинтеза. Нижняя граница данного слоя носит название компенсационного уровня. Выше этой глубины растения выделяют больше кислорода, чем потребляют, то избыточный кислород могут использовать другие организмы. Ниже этой глубины фотосинтез не может обеспечить дыхание, в связи с этим организмам доступен только кислород, который поступает с водой из более поверхностных слоев озера.
В светлых, поверхностных слоях воды обитают ярко и разнообразно окрашенные животные, глубоководные же виды обычно лишены пигментов. В сумеречной зоне океана обитают животные, окрашенные в цвета с красноватым оттенком, что помогает им скрываться от врагов, так как красный цвет в сине-фиолетовых лучах воспринимается как черный. Красная окраска характерна для таких животных сумеречной зоны, как морской окунь, красный коралл, различные ракообразные и др.
Поглощение света в воде тем сильнее, чем меньше ее прозрачность, что обусловлено наличием в ней частиц минеральных веществ (глина, ил). Уменьшается прозрачность воды и при бурном разрастании водной растительности в летний период или при массовом размножении мелких организмов, находящихся в поверхностных слоях во взвешенном состоянии. Прозрачность характеризуется предельной глубиной, где еще виден специально опускаемый диск Секки (белый диск диаметром 20 см). В Саргассовом море (самые прозрачные воды) диск Секки виден до глубины 66,5 м, в Тихом океане — до 59, в Индийском — до 50, в мелких морях — до 5-15 м. Прозрачность рек не превышает 1 -1,5 м, а в среднеазиатских реках Амударье и Сырдарье — нескольких сантиметров. Отсюда и границы зон фотосинтеза сильно колеблются в разных водоемах. В самых чистых водах зона фотосинтеза, или эуфотическая зона, достигает глубины не свыше 200 м, сумеречная (дисфотическая) простирается до 1000-1500 м, а глубже, в афотическую зону, солнечный свет совсем не проникает.
Световой день в воде значительно короче (особенно в глубоких слоях), чем на суше. Количество света в верхних слоях водоемов меняется и от широты местности, и от времени года. Так, длинные полярные ночи сильно ограничивают время, пригодное для фотосинтеза в арктических и приантарктических бассейнах, а ледовый покров затрудняет доступ света зимой во все замерзающие водоемы.
Солевой режим. В жизни водных организмов важную роль играет соленость воды или солевой режим. Химический состав вод формируется под влиянием естественно-исторических и геологических условий, а также при антропогенном воздействии. Содержание химических соединений (солей) в воде определяет ее соленость и выражается в граммах на литр или впромиле (°/од). По общей минерализации воды можно разделить на пресные с содержанием солей до 1 г/л, солоноватые (1-25 г/л), морской солености (26-50 г/л) и рассолы (более 50 г/л). Наиболее важными из растворенных веществ в воде являются карбонаты, сульфаты и хлориды.
Среди пресных вод много почти чистых, но много и таких, которые содержат до 0,5 г растворенных веществ на литр. Катионы по их содержанию в пресной воде располагаются следующим образом: кальций—64%, магний— 17%, натрий— 16%, калий — 3%. Это средние значения, а в каждом конкретном случае возможны колебания, иногда значительные.
Важным элементом в пресных водах является содержание кальция. Кальций может выступать в роли ограничивающего фактора. Различают воды «мягкие», бедные кальцием (менее 9 мг на 1 л), и воды «жесткие», содержание его в большом количестве (более 25 мг на 1 л).
В морской воде среднее содержание растворенных солей составляет 35 г/л, в окраинных морях значительно ниже. В морской воде обнаружены 13 металлоидов и не менее 40 металлов. По степени значимости первое место занимает поваренная соль, затем хлористый барий, сернокислый магний и хлористый калий.
Большинство водных битателей пойкилосмотичны. Осмотическое давление в их теле зависит от солености окружающей среды. Пресноводные животные и растения обитают в среде, где концентрация растворенных веществ ниже, чем в жидкостях тела и тканей. Из-за разницы в осмотическом давлении вне и внутри тела в организм постоянно проникает вода, вследствие чего гид-робионты пресных вод вынуждены интенсивно удалять ее. У них хорошо выражены процессы осморегуляции. У простейших это достигается работой выделительных вакуолей, у многоклеточных — удалением воды через выделительную систему. Некоторые инфузории каждые 2—2,5 мин выделяют количество воды, равное объему тела.
С повышением солености работа вакуолей замедляется, а при концентрации солей 17,5% перестает работать, так как разница осмотического давления между клетками и внешней средой исчезает.
Концентрация солей в жидкостях тела и тканей многих морских организмов изотонична концентрации растворенных солей в окружающей воде. В связи с этим осморегуляторные функции у них развиты слабее, чем у пресноводных. Осморегуляция является одной из причин того, что многие морские растения и животные не сумели заселить пресные водоемы и оказались типичными морскими жителями: кишечно-полостные (Coelenterata), иглокожие (Echinodermata), губки (Spongia), оболочники (Tunicata), погонофоры (Pogonophora). С другой стороны, в морях и океанах практически не обитают насекомые, тогда как пресноводные бассейны обильно ими заселены. Типично морские и типично пресноводные организмы не переносят значительных изменений солености и являются стеногалинными. Эвригалинных организмов, в частности животных, пресноводного и морского происхождения не так много. Они встречаются, нередко в больших количествах, в солоноватых водах. Это такие, как лещ (Abramis brama), пресноводный судак (Stizostedion lucioperca), щука (Ezox lucios), из морских — семейство кефалевых (Mugilidae).
Обитание растений в водной среде, помимо перечисленных выше особенностей, накладывает отпечаток и на другие стороны жизнедеятельности, особенно на водный режим у растений, в прямом смысле окруженных водой. У таких растений транспирации нет, а следовательно, и нет «верхнего двигателя», поддерживающего ток воды в растении. И вместе с тем ток, доставляющий к тканям питательные вещества, существует (правда, значительно слабее, чем у сухопутных растений), с ясно вьфаженной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению (у прикрепленных видов) и деятельности специальных клеток, выделяющих воду, — водяных устьиц или гидатод.
В пресных водах распространены растения, укрепленные на дне водоема. Часто их фотосинтетическая поверхность располагается над водой. К ним относятся камыши (Scirpus), кувшинки (Nymphaea), кубышки (Nyphar), рогозы (Typha), стрелолист (Sagittaria). У других фотосинтезирующие органы погружены в воду. Это рдесты (Potamogeton), уруть (Myriophyllum), элодея (Elodea). Отдельные виды высших растений пресных вод лишены корней и свободно плавают или обрастают подводные предметы, водоросли, которые прикреплены к грунту.
Газовый режим. Основными газами в водной среде являются кислород и углекислый газ. Остальные, такие, как сероводород или метан, имеют второстепенное значение.
Кислород для водной среды — важнейший экологический фактор. Он поступает в воду из воздуха и выделяется растениями при фотосинтезе. Коэффициент диффузии кислорода в воде примерно в 320 тыс. раз ниже, чем в воздухе, а общее его содержание в верхних слоях воды составляет 6—8 мл/л, или в 21 раз ниже, чем в атмосфере. Содержание кислорода в воде обратно пропорционально температуре. С повышением температуры и солености воды концентрация в ней кислорода понижается. В слоях, сильно заселенных животными и бактериями, может создаваться дефицит кислорода из-за усиленного его потребления. Так, в Мировом океане богатые жизнью глубины от 50 до 1000 м характеризуются резким ухудшением аэрации. Она в 7—10 раз ниже, чем в поверхностных водах, населенных фитопланктоном. Около дна водоемов условия могут быть близкими к анаэробным.
При застойном режиме в небольших водоемах вода также резко обедняется кислородом. Дефицит его может возникнуть и зимой подо льдом. При концентрации ниже 0,3—3,5 мл/л жизнь аэробов в воде невозможна. Содержание кислорода в условиях водоема оказывается лимитирующим фактором.
Среди водных обитателей значительно количество видов, способных переносить широкие колебания содержания кислорода в воде, близкие к его отсутствию. Это так называемыеэвриоксибионты. К ним относятся пресноводные олигохеты (Tubifex tubifex), брюхоногие моллюски (Viviparus viviparus). Очень слабое насыщение воды кислородом из рыб могут выдерживать сазан, линь, караси. Однако многие виды являются стеноксибионтными, т. е. могут существовать только при достаточно высоком насыщении воды кислородом, например радужная форель, кумжа, гольян и др. Многие виды живых организмов способны при недостатке кислорода впадать в неактивное состояние, так называемый аноксибиоз, и таким образом переживать неблагоприятный период.
Дыхание гидробионтов осуществляется как через поверхность тела, так и через специализированные органы — жабры, легкие, трахеи. Нередко покровы тела могут служить дополнительным органом дыхания. У отдельных видов встречается комбинирование водного и воздушного дыхания, например, двоякодышащие рыбы, сифонофоры, дискофанты, многие легочные моллюски, ракообразные Yammarus lacustris и др. Вторичноводные животные сохраняют обычно атмосферный тип дыхания как энергетически более выгодный, и поэтому нуждаются в контактах с воздушной средой. К ним относятся ластоногие, китообразные, водяные жуки, личинки комаров и т. д.
Углекислый газ. В водной среде живые организмы кроме недостатка света, кислорода могут испытывать недостаток доступной СО2, например растения для фотосинтеза. Углекислота поступает в воду в результате растворения СО2 содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. Содержание углекислого газа в воде колеблется в пределах 0,2—0,5 мл/л, или в 700 раз больше, чем в атмосфере. СО2 растворяется в воде в 35 раз лучше кислорода. Морская вода является главным резервуаром углекислого газа, так как содержит от 40 до 50 см3 газа на литр в свободной или связанной форме, что в 150 раз превышает его концентрацию в атмосфере.
Углекислый газ, содержащийся в воде, принимает участие в формировании известковых скелетных образований беспозвоночных животных и обеспечивает фотосинтез водных растений. При интенсивном фотосинтезе растений идет усиленное потребление углекислого газа (0,2—0,3 мл/л в час), что приводит к ее дефициту. На увеличение содержания СО2 в воде гидрофиты реагируют, повышая фотосинтез.
Дополнительным источником СО, для фотосинтеза водных растений является также углекислота, которая выделяется при разложении двууглекислых солей и их переходе в углекислые.
Малорастворимые карбонаты, которые при этом образуются, оседают на поверхность листьев в виде известкового налета или корочки, хорошо заметной при обсыхании многих водных растений.
Концентрация водородных ионов (рН) нередко сказывается на распределении водных организмов. Пресноводные бассейны с рН 3,7— 4,7 считаются кислыми, 6,95—7,3 нейтральными, с рН более 7,8 — щелочными. В пресных водоемах рН испытывает значительные колебания, нередко в течение суток. Морская вода более щелочная, и рН ее меньше изменяется, чем пресной. С глубиной рН уменьшается.
Из растений при рН меньше 7,5 растут полушник (Jsoetes), ежеголовник (Sparganium). В щелочной среде (рН 7,7—8,8) распространены многие виды рдестов, элодея, при рН 8,4—9 сильного развития достигает Typha angustifolia. Кислые воды торфяников способствуют развитию сфагновых мхов.
Большинство пресноводных рыб выдерживает рН от 5 до 9. Если рН меньше 5, наблюдается массовая гибель рыб, а выше 10 — погибают все рыбы и другие животные.
В озерах с кислой средой часто встречаются личинки двукрылых из рода Chaoborus, а в кислых водах болот распространены раковинные корненожки (Testaceae), отсутствуют пластинчато-жаберные моллюски из рода беззубок (Unio), редко встречаются другие моллюски.
Экологическая пластичность организмов водной среды. Вода является более стабильной средой, и абиотические факторы претерпевают сравнительно незначительные колебания, а отсюда водные организмы обладают по сравнению с наземными меньшей экологической пластичностью. Пресноводные растения и животные более пластичны, чем морские, так как пресная вода как среда жизни более изменчива. Оценивают широту экологической пластичности гидробионтов не только в целом к комплексу факторов (эври- и стенобионтность), но и по отдельности.
Так, установлено, что прибрежные растения и животные в отличие от обитателей открытых зон главным образом эвритермные и эвригалинные организмы, вследствие того, что температурные условия и солевой режим вблизи берега довольно изменчивы — прогреваемость солнцем и относительно интенсивное охлаждение, опреснение притоком воды из ручьев и рек, в частности в период дождей, и т. д. В качестве примера можно привести лотос, который относится к типичным стенотермным видам, произрастает только в мелких, хорошо прогреваемых водоемах. Обитатели поверхностных слоев по сравнению с глубоководными формами по вышеуказанным причинам оказываются более эвритермными и эвригалинными.
Экологическая пластичность является важным регулятором расселения организмов. Доказано, что гидробионты с высокой экологической пластичностью распространены широко, например, элодея. Противоположный же пример — рачок артемия (Artemia solina), живущий в небольших водоемах с очень соленой водой, является типичным стеногалинным представителем с узкой экологической пластичностью. По отношению же к другим факторам он обладает значительной пластичностью и в соленых водоемах встречается довольно часто.
Экологическая пластичность зависит от возраста и фазы развития организма. Например, морской брюхоногий моллюск Littorina во взрослом состоянии при отливах ежедневно длительное время обходится без воды, однако его личинки ведут планктонный образ жизни и не переносят высыхания.
Пожалуйста, ответьте на наши опросы. Несколько минут, потраченных вами на голосование, позволят нам понять, как сделать наш сайт и портал более интересным и нужным!


